Electroconvulsive therapy (ECT) remains an effective somatic treatment for a variety of psychiatric disorders. Despite its introduction almost a century ago, its site and mode of action remains elusive. One method of obtaining relevant information is by the recording of sensory evoked potentials from patients undergoing ECT. However, such human data is plagued with problems of methodology and interpretation. These limitations are not an impediment when employing an animal model of electroconvulsive shock (ECS). In the present experiment, the effects of ECS were studied on the superior colliculus visual evoked potential (SCVEP) in the non-medicated rat. The SCVEP was used as a measure of activity in the subcortical visual system as, in the rodent, the SC lies directly below the visual cortex. Immediately after the induction of generalised seizure activity (GSA) by ECS, all the components of the SCVEP were still preserved, basically intact. There was, however, a marked but very transient attenuation in its waveform not associated with any increase in the latency of the primary component. These findings are compared to a previous study where the cortical VEP was found to be completely abolished for up to two minutes following ECS. Judging solely by these and related neurophysiological data recorded from other exteroceptive sensory systems, it is concluded that the seat of ECT’s mode of action may lie quite discretely at the cortical level. Nevertheless, ECT has a profusion of effects on cerebral structure and function. Therefore, at the present time, this putative insight may be most applicable to an understanding of the negative or adverse (usually cognitive) side-effects which accompany ECT rather than to its positive (therapeutic) benefits. The acute loss of amplitude in the SCVEP waveform may not necessarily indicate that GSA impacts SC function, even momentarily. Instead, it is suggested that the interference more likely reflects a deficit in retinal processing which has been transferred to the SC.
| Published in | American Journal of Psychiatry and Neuroscience (Volume 13, Issue 2) |
| DOI | 10.11648/j.ajpn.20251302.13 |
| Page(s) | 61-71 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2025. Published by Science Publishing Group |
Electroconvulsive Shock, Electroconvulsive Therapy, Flash Visual Evoked Potential, Gap Junction, Generalized Seizure Activity, Superior Colliculus, Occipital Cortex
AEP | Auditory Evoked Potential |
BAEP | Brainstem Auditory Evoked Potential |
CAP | Compound Action Potential |
ECS | Electroconvulsive Shock |
ECT | Electroconvulsive Therapy |
EEG | Electroencephalogram |
EP | Evoked Potential |
ERG | Electroretinogram |
GSA | Generalized Seizure Activity |
PSP | Post Synaptic Potential |
SC | Superior Colliculus |
SCVEP | Superior Colliculus Visual Evoked Potential |
SEP | Somatosensory Evoked Potential |
VEP | Visual Evoked Potential |
| [1] | Kalinowsky, L. B., Hippius, H. Pharmacological, convulsive and other somatic treatments in psychiatry. New York: Grune and Stratton; 1969. |
| [2] | Sainsbury, M. J. Key to psychiatry. Sydney: ANZ Book Company, 1973. |
| [3] | Crowe, R. R. Electroconvulsive therapy- A current perspective. The New England Journal of Medicine. 1984, 311, 163-167. |
| [4] | Weiner, R. D. Does electroconvulsive therapy cause brain damage? The Behavioral and Brain Sciences. 1984, 7, 1-53. |
| [5] | Kaplan, H. I., Sadock, B. J., Grebb, J. A. Kaplan and Sadock’s synopsis of psychiatry: Behavioral sciences/ clinical psychiatry. 7th edition. Baltimore: Williams and Wilkins; 1994. |
| [6] | Bolwig, T. G. How does electroconvulsive therapy work? Theories on its mechanism. Canadian Journal of Psychiatry. 2011, 56. 13-18. |
| [7] | McCall, W. V., Andrade C., Sienaert, P. Searching for the mechanism(s) of ECT’s therapeutic effect. The Journal of ECT. 2014, 30, 87-89. |
| [8] | Sienaert, P. Mechanisms of ECT: Reviewing the science and dismissing the myths. The Journal of ECT. 2014, 30, 85-86. |
| [9] | Sackeim, H. A. Modern electroconvulsive therapy: vastly improved yet greatly underused. JAMA Psychiatry. 2017, 74, 779-780. |
| [10] | Singh, A., Kar, S. K. How electroconvulsive therapy works? understanding the neurobiological mechanisms. Clinical Psychopharmacology and Neuroscience. 2017, 15, 210. |
| [11] | Kirov, G., Jauhar, S., Sienaert, P., Kellner, C, H., Mc Loughlin, D. M. Electroconvulsive therapy for depression: 80 years of progress. The British Journal of Psychiatry. 2021, 219, 594-597. |
| [12] | Tor, P. C., Tan, X. W., Martin, D., Loo, C. Comparative outcomes in electroconvulsive therapy (ECT): A naturalistic comparison between outcomes in psychosis, mania, depression, psychotic depression and catatonia. European Neuropsychopharmacology. 2021, 51, 43 – 54. |
| [13] | Potter, W. Z., Rudorfer, M. V. Electroconvulsive therapy – A modern medical procedure. The New England Journal of Medicine. 1993, 328, 882-883. |
| [14] | Espinoza, R. T., Kellner, C. H. Electroconvulsive therapy. The New England Journal of Medicine. 2022, 386, 667-672. |
| [15] | Takamiya, A., Seki, M., Kudo, S., Yoshizaki, T., Nakahara, J., Mimura, M., Kishimoto, T. Electroconvulsive therapy for Parkinson’s disease: A systematic review and meta-analysis. Movement Disorders. 2021, 36, 50-58. |
| [16] | Small, J. G., Small, I. F., Milstein, V. Electrophysiology of EST. In Psychopharmacology: A Generation of Progress, Lipton, M. A., DiMascio, A., Killam, K. F., Eds., New York: Raven Press; 1978, pp. 759-769. |
| [17] | Pinel, J. P. J. After forty-five years ECT is still controversial. The Behavioral and Brain Sciences. 1984, 7, 30-31. |
| [18] | Sackeim, H. A. Central issues regarding the mechanisms of action of electroconvulsive therapy: Directions for future research. Psychopharmacology Bulletin. 1994, 30, 281-308. |
| [19] | Fink, M. Convulsive therapy: theory and practice. New York: Raven Press; 1979. |
| [20] | Small, J. G., Small, I. F., Perez, H. C., Sharpley, P. Electroencephalographic and neurophysiological studies of electrically induced seizures. The Journal of Nervous and Mental Disease. 1970, 150, 479-489. |
| [21] | Small, I. F., Small, J. G. Electroencephalographic (EEG), evoked potential, and direct current (DC) responses with unilateral electroconvulsive treatment (ECT). The Journal of Nervous and Mental Disease. 1971, 152, 396-404. |
| [22] | Kriss, A., Halliday, A. M., Halliday, E., Pratt, R. T. C. Evoked potentials following unilateral ECT. I. The somatosensory evoked potential. Electroencephalography and Clinical Neurophysiology. 1980, 48, 481-489. |
| [23] | Kriss, A., Halliday, A. M., Halliday, E., Pratt, R. T. C. Evoked potentials following unilateral ECT. II. The flash evoked potential. Electroencephalography and Clinical Neurophysiology. 1980, 48, 490-501. |
| [24] | Small, J. G., Milstein, V., Kellams, J. J., Small, I. F. Auditory brainstem evoked responses in hospitalized patients undergoing drug treatment or ECT. Biological Psychiatry. 1981, 16, 287-290. |
| [25] | Weiner, R. D., Erwin, C. W., Weber, B. A. Acute effects of electroconvulsive therapy on brain stem auditory- evoked potentials. Electroencephalography and Clinical Neurophysiology. 1981, 52, 202-204. |
| [26] | Kolbeinsson, H., Petursson, H. Electroencephalographic correlates of electroconvulsive therapy. Acta Psychiatrica Scandinavica. 1988, 78, 162-168. |
| [27] | Shaw, N. A. Effect of electroconvulsive shock on the somatosensory evoked potential in the rat. Experimental Neurology. 1985, 90, 566-579. |
| [28] | Shaw, N. A. Effect of electroconvulsive shock on the cervical evoked potential in the rat. Experimental Neurology. 1986, 91, 646-649. |
| [29] | Shaw, N. A. The effect of electroconvulsive shock on the brain stem auditory evoked potential in the rat. Biological Psychiatry. 1986, 21, 1327-1331. |
| [30] | Shaw, N. A. Effect of electroconvulsive shock on the slow components of the brain stem auditory evoked potential. Experimental Neurology. 1988, 100, 242-247. |
| [31] | Shaw, N. A. The effects of generalized seizure activity on the cerebellar auditory potential. International Journal of Neuroscience. 1995, 83, 17-23. |
| [32] | Shaw, N. A. The effects of electroconvulsive shock on the collicular auditory potential. Biological Psychiatry. 1996, 39, 223-226. |
| [33] | Shaw, N. A. The effects of electroconvulsive shock on the flash visual evoked potential in the rat. Electroencephalography and Clinical Neurophysiology. 1997, 104, 180-187. |
| [34] | Shaw, N. A. The effects of electroconvulsive shock on the thalamic auditory potential. Biological Psychiatry. 1996, 40, 807-810. |
| [35] | Shaw, N. A. The effects of electroconvulsive shock on the primary cortical auditory potential. Biological Psychiatry. 1998, 43, 595-600. |
| [36] | Shaw, N. A. The effects of electroconvulsive shock on the short-latency somatosensory evoked potential in the rat. Brain Research Bulletin. 1998, 45, 427-433. |
| [37] | Shaw, N. A. The effects of electroconvulsive shock on retinal activity. Physiology and Behavior. 1999, 67, 153-159. |
| [38] | Polyak, S. L. The vertebrate visual system. Chicago: University of Chicago Press; 1957. |
| [39] | Thompson, R. A behavioral atlas of the rat brain. New York: Oxford University Press; 1978. |
| [40] | Smith, C. G. Basic neuroanatomy. 2nd edition. Toronto: University of Toronto Press; 1972. |
| [41] | Sefton, A. J., Dreher, B. Visual system. In The Rat Nervous System, Volume 1, Paxinos, G., Ed., Sydney: Academic Press; 1985, pp. 169-221. |
| [42] | Xintaras, C., Johnson, B. L., Ulrich, C. E. Terrill, R. E., Sobecki, M. F. Application of the evoked response technique in air pollution toxicology. Toxicology and Applied Pharmacology. 1966, 8, 77-87. |
| [43] | Woolley, D. E. Some aspects of neurophysiological basis of insecticide action. Federation Proceedings. 1976, 35, 2610-2617. |
| [44] | Dyer, R. S., Annau, Z. Carbon monoxide and flash evoked potentials from rat cortex and superior colliculus. Pharmacology, Biochemistry, and Behavior. 1977, 6, 461-465. |
| [45] | Hetzler, B. E., Oaklay, K. E. Dose effects of pentobarbital on evoked potentials in visual cortex and superior colliculus of the albino rat. Neuropharmacology. 1981, 20, 969-978. |
| [46] | Hetzler, B. E., Heilbronner, R. L., Griffin, J., Griffin, G. Acute effects of alcohol on evoked potentials in visual cortex and superior colliculus of the rat. Electroencephalography and Clinical Neurophysiology. 1981, 51, 69-79. |
| [47] | Hetzler, B. E., Oaklay, K. E., Heilbronner, R. L., Vestal, T. Acute effects of alcohol on photic evoked potentials of albino rats: visual cortex and superior colliculus. Pharmacology, Biochemistry, and Behavior. 1982, 17, 1313-1316. |
| [48] | Hetzler B. E., Berger, L. K. Ketamine-induced modification of photic evoked potentials in the superior colliculus of hooded rats. Neuropharmacology. 1984, 23, 473-476. |
| [49] | Hetzler, B. E., Melk, A. M. Effects of ketamine, naloxone, and physostigmine on flash evoked potentials in rat superior colliculus. Pharmacology, Biochemistry, and Behavior. 1989, 32, 511-518. |
| [50] | Hetzler, B. E., Zeisset, H. L. THIP, A selective gamma-aminobutyric acid receptor agonist, alters flash-evoked potentials in rats. Pharmacology, Biochemistry, and Behavior. 1997, 58, 923-932. |
| [51] | Hetzler, B. E., Burkard, H. K. Effects of dizocilpine (MK-801) on flash – evoked potentials, body temperature, and locomotor activity of hooded rats. Pharmacology, Biochemistry, and Behavior. 1999, 62, 559-573. |
| [52] | Hetzler, B. E., Theinpeng, M. Nicotine alters flash-evoked potentials in Long-Evans rats. Pharmacology, Biochemistry, and Behavior. 2004, 77, 717-729. |
| [53] | Hetzler, B. E., Ondracek, J. M. Baclofen alters flash-evoked potentials in Long-Evans rats. Pharmacology, Biochemistry, and Behavior. 2007, 86, 727-740. |
| [54] | Gowan J. D., Coizet, V., Devonshire, I. M., Overton, P. G. D-amphetamine depresses visual responses in the rat superior colliculus: a possible mechanism for amphetamine-induced decreases in distractibility. Journal of Neural Transmission. 2008, 115, 377-387. |
| [55] | Hetzler, B. E., Beckman, E. J. Effects of mecamylamine on flash – evoked potentials, body temperature, and behavior in Long-Evans rats. Pharmacology, Biochemistry, and Behavior. 2011, 97, 660-668. |
| [56] | Hetzler, B. E., Meckel, K. R., Stickle, B. A. Methylphenidate alters flash-evoked potentials, body temperature, and behavior in Long-Evans rats. Pharmacology, Biochemistry, and Behavior. 2014, 116, 76-83. |
| [57] | Dyer, R. S., Annau, Z. Flash evoked potentials from rat superior colliculus. Pharmacology, Biochemistry, and Behavior. 1977, 6, 453-459. |
| [58] | Miller, N. E., Dicara L. Instrumental learning of heart rate changes in curarized rats: shaping and specificity to discriminative stimulus. Journal of Comparative and Physiological Psychology. 1967, 63, 12-19. |
| [59] | Smith, S. M., Brown, H. O., Toman, J. E. P., Goodman, L. S. The lack of cerebral effects of d – tubocurarine. Anesthesiology. 1947, 8, 1-14. |
| [60] | Whitham, E. M., Pope, K. J., Fitzgibbon, S. P., Lewis, T., Clark, C. R., Loveless, S., Broberg, M., Wallace, A., DeLosAngeles, D., Lillie, P., Hardy, A., Fronsko, R., Pulbrook, A., Willoughby, J. O. Scalp electrical recording during paralysis: quantitative evidence that EEG frequencies above 20 Hz are contaminated by EMG. Clinical Neurophysiology. 2007, 118, 1877-1888. |
| [61] | Trowill, J. A. Instrumental conditioning of the heart rate in the curarized rat. Journal of Comparative and Physiological Psychology. 1967, 63, 7-11. |
| [62] | Shavalia, D. A., Dodge, A. M., Beatty, W. W. Time – dependent effects of ECS on spatial memory in rats. Behavioral and Neural Biology. 1981, 31, 261-273. |
| [63] | Achor, L. J., Starr, A. Auditory brain stem responses in the cat. II. Effects of lesions. Electroencephalography and Clinical Neurophysiology. 1980, 48, 174-190. |
| [64] | Foutz, A. S., Dauthier, C., Kerdelhue, B. β – Endorphin plasma levels during neuromuscular blockade in unanesthetized cat. Brain Research. 1983, 263, 119-123. |
| [65] | Hetzler, B. E., Martin, E. I. Nicotine-ethanol interactions in flash-evoked potentials and behavior of Long-Evans rats. Pharmacology, Biochemistry, and Behavior. 2006, 83, 76-89. |
| [66] | Sefton, A. J. The electrical activity of the anterior colliculus in the rat. Vision Research. 1969, 9, 207-222. |
| [67] | Goldberg, M. E., Robinson, D. L. Visual system: superior colliculus. In Handbook of Behavioral Neurobiology, Volume 1: Sensory Integration, Masterton, R. B., Ed., New York: Plenum Press; 1978, pp. 119 – 164. |
| [68] | Brodal, A. Neurological anatomy. 3rd edition. New York: Oxford University Press; 1981. |
| [69] | Shaw, N. A. The relationship between the putative optic pathway potential and the electroretinogram. Physiology and Behavior. 1998, 63, 615 – 620. |
| [70] | Wood, C. C., Allison, T. Interpretation of evoked potentials: A neurophysiological perspective. Canadian Journal of Psychology. 1981, 35, 113-135. |
| [71] | Greenberg, R. P., Ducker, T. B. Evoked potentials in the clinical neurosciences. Journal of Neurosurgery. 1982, 56, 1- 18. |
| [72] | Kuffler, S. W., Nicholls, J. G. From neuron to brain. Sunderland: Sinauer; 1976. |
| [73] | Schmidt, F. O., Dev, P., Smith, B. H. Electrotonic processing of information by brain cells. Science. 1976, 193, 114-120. |
| [74] | Ottoson, D. Physiology of the nervous system. London: MacMillan; 1983. |
| [75] | Hall, Z. W. An introduction to molecular neurobiology. Sunderland: Sinauer; 1992. |
| [76] | Kandel, E. R., Siegelbaum, S. A. Overview of synaptic transmission. In Principles of Neural Science. 4th edition, Kandel, E. R., Schwartz, J. H., Jessell, T. M., Eds., New York: McGraw-Hill; 2000, pp. 175 – 186. |
| [77] | Connors, B. W., Long, M. A. Electrical synapses in the mammalian brain. Annual Review of Neuroscience. 2004, 27, 393-418. |
| [78] | Sohl, G., Maxeiner, S., Willecke, K. Expression and functions of neuronal gap junctions. Nature Reviews Neuroscience. 2005, 6, 191-200. |
| [79] | Vaughn, M. J., Haas, J. S. On the diverse functions of electrical synapses. Frontiers in Cellular Neuroscience. 2022, 16, 910015. |
| [80] | Leonard, B. E. Fundamentals of psychopharmacology. 2nd edition. Chichester: Wiley; 1997. |
| [81] | Madsen, T. M., Treschow, A., Bengzon, J., Bolwig, T. G., Lindvall, O., Tingstrom, A. Increased neurogenesis in a model of electroconvulsive therapy. Biological Psychiatry. 2000, 47, 1043 – 1049. |
| [82] | Merkl, A., Heuser, I., Bajbouj, M. Antidepressant electroconvulsive therapy: mechanism of action, recent advances and limitations. Experimental Neurology. 2009, 219, 20 – 26. |
| [83] | Baldinger, P., Lotan, A., Frey, R., Kasper, S., Lever, B., Lanzenberger, R. Neurotransmitters and electroconvulsive therapy. The Journal of ECT. 2014, 30, 116 – 121. |
| [84] | Bouckaert, F., Sienaert, P., Obbels, J., Dols, A., Vandenbulcke, M., Stek, M., Bolwig, T. ECT: its brain enabling effects: a review of electroconvulsive therapy – induced structural brain plasticity. The Journal of ECT. 2014, 30, 143–151. |
| [85] | Cano, M., Camprodon, J. A. Understanding the mechanisms of action of electroconvulsive therapy: Revisiting neuroinflammatory and neuroplasticity hypotheses. JAMA Psychiatry. 2023, 80, 643 – 644. |
| [86] | Carney, M. W. P., Sheffield, B. F. Electroconvulsive therapy and the diencephalon. The Lancet. 1973, 1505–1506. |
| [87] | Abrams, R., Taylor, M. A. Electroconvulsive therapy and the diencephalon: A preliminary report. Comprehensive Psychiatry. 1974, 15, 233 – 236. |
| [88] | Abrams, R., Taylor, M. A. Diencephalic stimulation and the effects of ECT in endogenous depression. British Journal of Psychiatry. 1976, 129, 482–485. |
| [89] | Fink, M. A neuroendocrine theory of convulsive therapy. Trends in Neurosciences. 1980, 3, 25–27. |
| [90] | Fink, M., Ottosson, J-O. A theory of convulsive therapy in endogenous depression: Significance of hypothalamic functions. Psychiatry Research. 1980, 2, 49–61. |
| [91] | Haskett, R. Electroconvulsive therapy’s mechanism of action: Neuroendocrine hypotheses. The Journal of ECT. 2014, 30, 107–110. |
| [92] | Semkovska, M., McLoughlin, D. M. Objective cognitive performance associated with electroconvulsive therapy for depression: a systematic review and meta-analysis. Biological Psychiatry. 2010, 68, 568–577. |
| [93] | Altman, J., Malis, L. I. An electrophysiological study of the superior colliculus and visual cortex. Experimental Neurology. 1962, 5, 233–249. |
APA Style
Shaw, N. A. (2025). The Effects of Electroconvulsive Shock on the Superior Colliculus Visual Evoked Potential. American Journal of Psychiatry and Neuroscience, 13(2), 61-71. https://doi.org/10.11648/j.ajpn.20251302.13
ACS Style
Shaw, N. A. The Effects of Electroconvulsive Shock on the Superior Colliculus Visual Evoked Potential. Am. J. Psychiatry Neurosci. 2025, 13(2), 61-71. doi: 10.11648/j.ajpn.20251302.13
AMA Style
Shaw NA. The Effects of Electroconvulsive Shock on the Superior Colliculus Visual Evoked Potential. Am J Psychiatry Neurosci. 2025;13(2):61-71. doi: 10.11648/j.ajpn.20251302.13
@article{10.11648/j.ajpn.20251302.13,
author = {Nigel Alexander Shaw},
title = {The Effects of Electroconvulsive Shock on the Superior Colliculus Visual Evoked Potential
},
journal = {American Journal of Psychiatry and Neuroscience},
volume = {13},
number = {2},
pages = {61-71},
doi = {10.11648/j.ajpn.20251302.13},
url = {https://doi.org/10.11648/j.ajpn.20251302.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajpn.20251302.13},
abstract = {Electroconvulsive therapy (ECT) remains an effective somatic treatment for a variety of psychiatric disorders. Despite its introduction almost a century ago, its site and mode of action remains elusive. One method of obtaining relevant information is by the recording of sensory evoked potentials from patients undergoing ECT. However, such human data is plagued with problems of methodology and interpretation. These limitations are not an impediment when employing an animal model of electroconvulsive shock (ECS). In the present experiment, the effects of ECS were studied on the superior colliculus visual evoked potential (SCVEP) in the non-medicated rat. The SCVEP was used as a measure of activity in the subcortical visual system as, in the rodent, the SC lies directly below the visual cortex. Immediately after the induction of generalised seizure activity (GSA) by ECS, all the components of the SCVEP were still preserved, basically intact. There was, however, a marked but very transient attenuation in its waveform not associated with any increase in the latency of the primary component. These findings are compared to a previous study where the cortical VEP was found to be completely abolished for up to two minutes following ECS. Judging solely by these and related neurophysiological data recorded from other exteroceptive sensory systems, it is concluded that the seat of ECT’s mode of action may lie quite discretely at the cortical level. Nevertheless, ECT has a profusion of effects on cerebral structure and function. Therefore, at the present time, this putative insight may be most applicable to an understanding of the negative or adverse (usually cognitive) side-effects which accompany ECT rather than to its positive (therapeutic) benefits. The acute loss of amplitude in the SCVEP waveform may not necessarily indicate that GSA impacts SC function, even momentarily. Instead, it is suggested that the interference more likely reflects a deficit in retinal processing which has been transferred to the SC.
},
year = {2025}
}
TY - JOUR T1 - The Effects of Electroconvulsive Shock on the Superior Colliculus Visual Evoked Potential AU - Nigel Alexander Shaw Y1 - 2025/06/16 PY - 2025 N1 - https://doi.org/10.11648/j.ajpn.20251302.13 DO - 10.11648/j.ajpn.20251302.13 T2 - American Journal of Psychiatry and Neuroscience JF - American Journal of Psychiatry and Neuroscience JO - American Journal of Psychiatry and Neuroscience SP - 61 EP - 71 PB - Science Publishing Group SN - 2330-426X UR - https://doi.org/10.11648/j.ajpn.20251302.13 AB - Electroconvulsive therapy (ECT) remains an effective somatic treatment for a variety of psychiatric disorders. Despite its introduction almost a century ago, its site and mode of action remains elusive. One method of obtaining relevant information is by the recording of sensory evoked potentials from patients undergoing ECT. However, such human data is plagued with problems of methodology and interpretation. These limitations are not an impediment when employing an animal model of electroconvulsive shock (ECS). In the present experiment, the effects of ECS were studied on the superior colliculus visual evoked potential (SCVEP) in the non-medicated rat. The SCVEP was used as a measure of activity in the subcortical visual system as, in the rodent, the SC lies directly below the visual cortex. Immediately after the induction of generalised seizure activity (GSA) by ECS, all the components of the SCVEP were still preserved, basically intact. There was, however, a marked but very transient attenuation in its waveform not associated with any increase in the latency of the primary component. These findings are compared to a previous study where the cortical VEP was found to be completely abolished for up to two minutes following ECS. Judging solely by these and related neurophysiological data recorded from other exteroceptive sensory systems, it is concluded that the seat of ECT’s mode of action may lie quite discretely at the cortical level. Nevertheless, ECT has a profusion of effects on cerebral structure and function. Therefore, at the present time, this putative insight may be most applicable to an understanding of the negative or adverse (usually cognitive) side-effects which accompany ECT rather than to its positive (therapeutic) benefits. The acute loss of amplitude in the SCVEP waveform may not necessarily indicate that GSA impacts SC function, even momentarily. Instead, it is suggested that the interference more likely reflects a deficit in retinal processing which has been transferred to the SC. VL - 13 IS - 2 ER -
Department of Anatomy and Medical Imaging, University of Auckland, Auckland, New Zealand
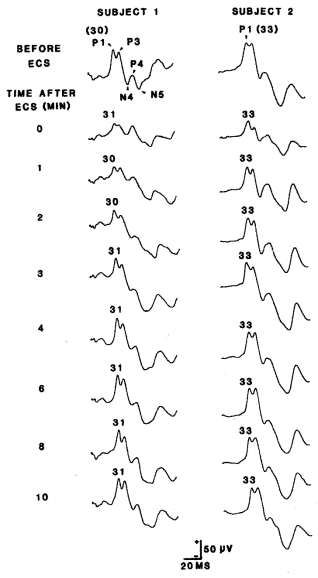
Figure 1. Two examples of the effects of ECS on the SCVEP. In the baseline examples, the primary collicullar component (P1) is identified with its actual latency (ms) in parentheses. In the post – ECS SCVEPs, only the latency of P1 is indicated. SCVEPs recorded at 5, 7, and 9 minutes are not illustrated. In the baseline recording of subject 1, all 5 principal components of the waveform are identified.
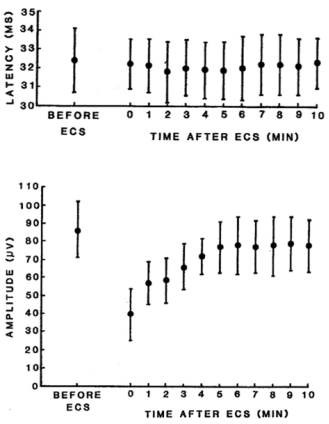
Figure 2. Mean latency and amplitude (± 1 SD) of the P1 component of the SCVEP at the times indicated following ECS. The amplitude of P1 was calculated by referring it to the shallow negative trough preceding P1.
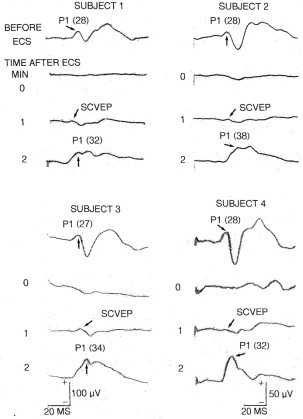
Figure 3. Four examples of the effects of ECS on the cortical VEP. The primary cortical response (P1) is indicated with the arrows and the actual latency is in parentheses. Note the absence of the waveform at 0 minutes but the appearance of a low amplitude positivity at 1 minute. This is a far field recording of the SCVEP, not the cortical potential. Note also the enhanced amplitude of P1 in the post-ictal recording at 2 minutes. Details on how these EPs were recorded are available elsewhere [33]. Illustrations of the waveforms were made using a photochemical process and in the interim the quality of them deteriorated and faded making reproduction less than optimal. In some traces (especially subject 1), either the beginning or end is missing but the latency values are accurate. The scale at the bottom left applies to both subjects 1 and 3 while the bottom right scale applies to subjects 2 and 4.

